Conformational cycle and ion-coupling mechanism of the Na+/hydantoin transporter Mhp1
By Kelli Kazmier, Shruti Sharma, Shahidul M Islam, Benoît Roux, and Hassane Mchaourab.
Published in Proc Natl Acad Sci U S A. 2014 Oct 14;111(41):14752-7. PMID: 25267652. PMCID: PMC4205665. Link to publication page.
Project: The Transport Cycle in Neurotransmitter Uptake Systems. Core Facility: Computational Modeling.
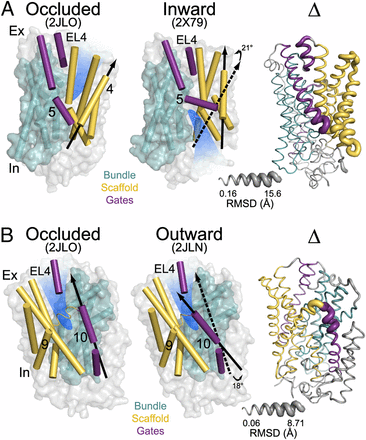
Figure 1. (A) Comparison of outward-facing (2JL0) and inward-facing crystal structures (2X79) of Mhp1 highlighting the inferred movement of the scaffold (yellow) relative to the bundle (cyan). The rmsd between the two structures is displayed by the ribbon thickness (scale in gray) on the 2JLO backbone (Δ). The change in orientation of the scaffold between the two structures is shown by the arrows. The approximate location of the solvent accessible vestibule is indicated in blue. (B) Comparison of outward-facing conformation (2JLO) and outward-facing/occluded conformation (2JLN) highlighting the inferred movement of TM10.
Significance
Na+-coupled symporters use the cellular Na+ gradient to power transport of physiologically important molecules across the lipid membrane. However, the mechanism by which binding and dissociation of Na+ drive transport remains undefined. This work investigated the Na+/hydantoin transporter Mhp1, a member of the LeuT-fold class of transporters, to describe the conformations sampled during its transport cycle and elucidate the ligand-induced shifts in its conformational equilibrium. The results of this study suggest that Mhp1 isomerization between inward- and outward-facing conformations are Na+-independent and that coupling to the Na+ gradient occurs through modulation of substrate affinity by Na+ coordination. A previously unidentified model of Mhp1 transport defined by ligand-independent equilibrium fluctuations emerges from this work, offering a new perspective on Na+-coupled symport in the LeuT-fold.
Abstract
Ion-dependent transporters of the LeuT-fold couple the uptake of physiologically essential molecules to transmembrane ion gradients. Defined by a conserved 5-helix inverted repeat that encodes common principles of ion and substrate binding, the LeuT-fold has been captured in outward-facing, occluded, and inward-facing conformations. However, fundamental questions relating to the structural basis of alternating access and coupling to ion gradients remain unanswered. Here, we used distance measurements between pairs of spin labels to define the conformational cycle of the Na+-coupled hydantoin symporter Mhp1 from Microbacterium liquefaciens. Our results reveal that the inward-facing and outward-facing Mhp1 crystal structures represent sampled intermediate states in solution. Here, we provide a mechanistic context for these structures, mapping them into a model of transport based on ion- and substrate-dependent conformational equilibria. In contrast to the Na+/leucine transporter LeuT, our results suggest that Na+ binding at the conserved second Na+ binding site does not change the energetics of the inward- and outward-facing conformations of Mhp1. Comparative analysis of ligand-dependent alternating access in LeuT and Mhp1 lead us to propose that different coupling schemes to ion gradients may define distinct conformational mechanisms within the LeuT-fold class.
 versus distance (r) between spin labels and reporting the ligand-dependent movement of the scaffold relative to the bundle on the intracellular (A) and extracellular (B) sides. The locations of the spin-label pairs are highlighted by black spheres connected by a line. Distance distributions for each pair were obtained in the apo, Na+-bound (Na+), and Na+- and BH-bound (Na+/BH) intermediates.")

 Distributions monitoring the distance from TM10 to bundle helices are consistent with a dynamic TM10 but do not reveal a change in its overall disposition as predicted from the occluded crystal structure. (B) Distance distributions between TM9 and helices in the scaffold (TM3) and bundle (TM9) have a broader width upon substrate binding, suggesting increased dynamic of TM9 and deviation from rigid-body movement in the scaffold. (C) Gating of the extracellular vestibule by EL4 is revealed by the substrate-dependent changes in distance distributions to the scaffold and the bundle. The asterisk denotes distance components arising from a minor aggregated population of Mhp1.")
, through low probability transitions, samples outward-facing conformations (priming, B), allowing the simultaneous binding of the Na+ and substrate (loading), and resulting in a stabilization of the outward-facing conformation (C). Low-probability fluctuations allow sampling of the inward-facing conformation (switch, D) where, driven by its concentration gradient, Na+ dissociates to the intracellular solution (release). In the absence of bound Na+, BH affinity to Mhp1 is reduced, which drives dissociation of BH to the intracellular side (E). The cycle continues through the isomerization of apo Mhp1 from inward-facing (A) to outward-facing (B). Show Less")
